|
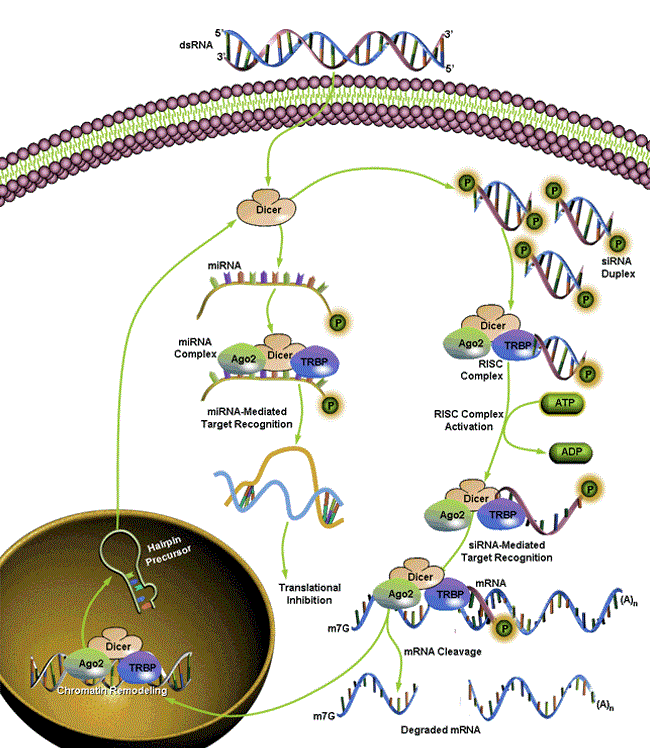
Short-interfering RNAs or siRNA suppress gene expression through a highly regulated enzyme-mediated process called RNA interference [2]. It was initially identified as a conserved surveillance system that occurs in a broad range of eukaryotic organisms [3]. RNAi can be initiated by exposing cells to dsRNA either via transfection or endogenous expression of the siRNA expression constructs as achieved by PCR [4]. RNAi involves multiple RNA-protein interactions characterized by four major steps; which is fundamentally the same in almost all the eukaryotes -
The assembly of RISCs can be traced from the biogenesis of the small RNA molecules and the recruitment of these RNAs by the RISC loading complex (RLC) [6]. dsRNA and proteins of the RNAi machinery can direct epigenetic alterations to homologous DNA sequences to induce transcriptional gene silencing or, in extreme cases, DNA elimination, providing direct silencing of certain gene. Interactions with RISC dictate siRNA localization even when siRNA is conjugated [7]. Another possible explanation for the conservation of the siRNA pathway is that many of the enzymes and protein factors that participate in this pathway are identical and structurally and functionally very similar to enzymes and protein factors that participate in a regulation pathway, which uses endogenous imperfect hairpin RNA structures called microRNAs or miRNAs that regulate the mRNA translation [8]. Imaging of transfer and accumulation of siRNA in cells can be achieved by high-resolution magnetic resonance imaging (MRI) and near-infrared in vivo optical imaging (NIRF) [10].
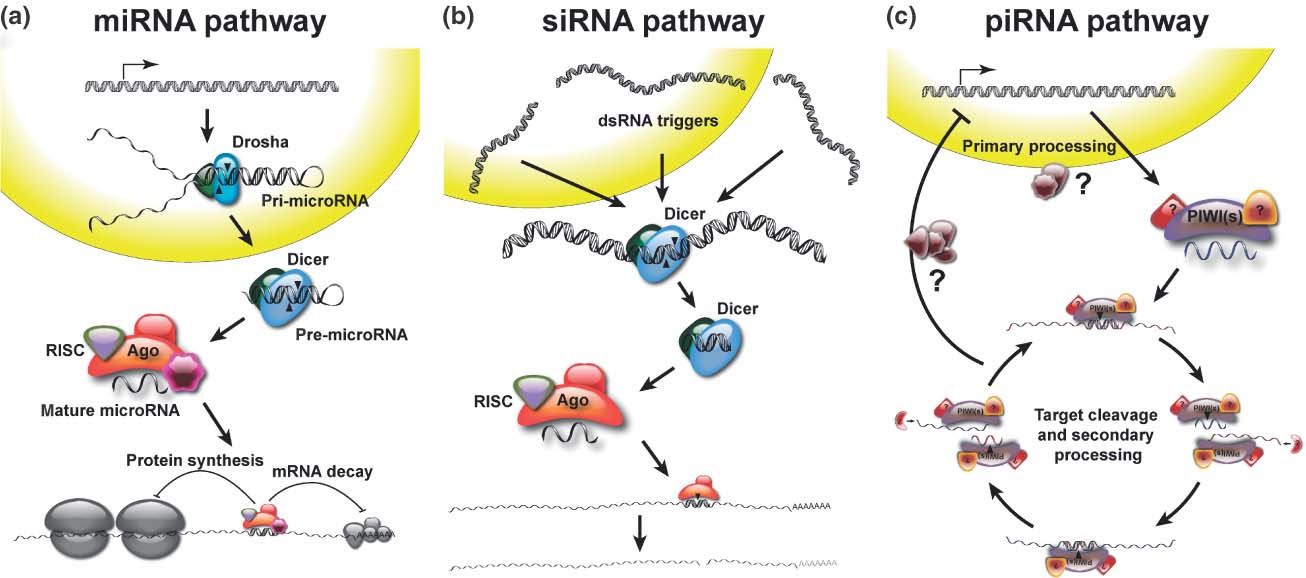
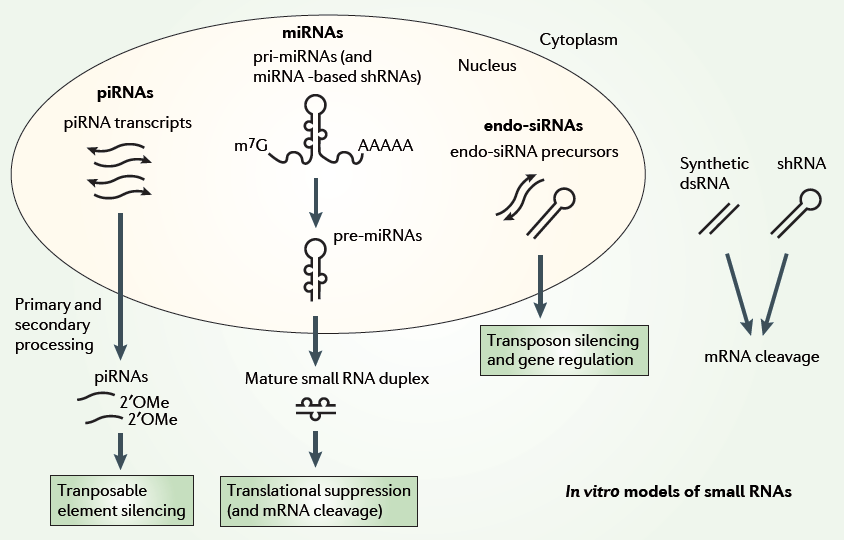
Biogenesis and Regulatory Features of miRNAs, siRNAs and piRNAs MicroRNA (miRNA) genes are transcribed by RNA polymerases II and III into primary transcripts called the pri-miRNAs. These are processed into pre-miRNAs in the nucleus by a Microprocessor complex, which contains the RNase III enzyme Drosha and the double-stranded RNA-binding protein DiGeorge syndrome critical region gene 8 (DGCR8; also known as Pasha). Some miRNAs, called mirtrons, have also been shown to be generated from introns that bypass Drosha requirement. Pre-miRNAs are then transported by exportin 5, which is a karyopherin, and RanGTP into the cytoplasm, where they are further processed by the RNAse III enzyme Dicer. This results in double-stranded 20–25 nucleotide (nt) intermediates with 2 nt overhangs on the 3' end. One of the RNA strands is then loaded by Dicer into an RNA-induced silencing complex (RISC), which contains an Argonaute-subfamily member, that then targets the 3' untranslated region of the target mRNAs by an imperfect match between the miRNA and the mRNA, to repress translation. However, it has been recently shown that miRNAs can activate translation of target mRNAs [46]. The small interfering RNA (siRNA) pathway initiates with either bidirectional transcription or transcription of an inverted repeat that results in a double-stranded precursor. This precursor is also processed in the cytoplasm by Dicer and is loaded into a small interfering RISC (siRISC) complex intended for target mRNA degradation [52].
The Piwi-interacting RNA (piRNA) pathway seems to be particularly active in the germ line and functions in transposon silencing and epigenetic regulation. The exact mechanism of biogenesis is still unclear. Presumably, piRNAs originate from a long single-stranded RNA polymerase transcript (1–100 kilobases) that is often transcribed from a bidirectional piRNA cluster (see the figure, panel c). The precursor is subsequently processed by unknown mechanisms, which probably involve Piwi proteins but not Dicer, into mature piRNAs of 23–32 nt in length[51]. Downloads | Glossary | References | Contact © 2012, Saie Mogre. All Rights Reserved. |
{kind=link}
{kind=link}