|
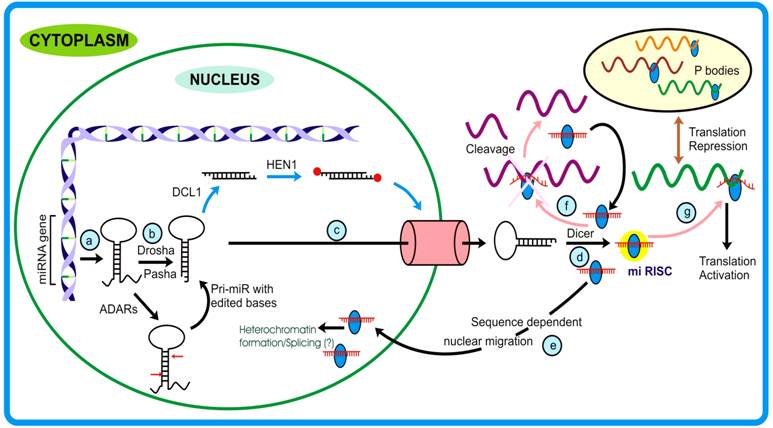
The discovery of microRNA dates back to 1993, when Lee et al. elucidated the function of a non-coding transcript in C. elegans, the expression of which varied spatio-temporally and the mutants showed developmental abnormalities. These genes were later recognized as precursor molecules of yet another important class of endogenous small RNA viz. microRNA that, unlike siRNAs, target messages different from that of the parent. MicroRNA genes constitute ~1% of the total coding genes and form the largest class of regulatory molecules [23]. MicroRNA biogenesisMicro RNA biogenesis is now believed to be operative by more than one pathway, as described below: Canonical miRNA pathway: MiRNAs are ~19-23 nt long single-stranded RNAs generated from single-stranded transcript having local-hairpin structure. These transcripts are generated by pol II and therefore possess 5′ cap and 3′ poly-A tail hallmarks. Unlike the maturation of large RNA classes that occurs in the nucleus, miRNA maturation begins in the nucleus and terminates in the cytoplasm. In animals, the nuclear processing initiates with the endonucleolytic activity of Drosha, an RNase III enzyme, which in association with Pasha, recognizes and generates the stem- loop structure (pre-miR) from the pri-miRs mediated by Exp-5 and Ran-GTPase, pre-miR are transported out to cytoplasm where these are acted upon by Dicer. Except for the different proteins that participate in miRNA pathway, plants follow almost similar miRNA biogenesis. However, an important difference between the miRNA biogenesis pathways of plants and animals lies in the fact that in plants the DCL1 acts on the pre-miRs in the nucleus [35]. In Arabidopsis and Drosophila, the duplex small RNAs are acted upon by methyl transferases that add methyl group at the 2′-hydroxyl residues of the terminal ribose sugars. Such end-modification protects these RNA species from any kind of degradation or uridylation [36, 38]. Duplex miRs are then unwound by Argonaute1, a prominent member of miRNA- RISC assembly (miRNP), generating mature miR. The mi-RISC/ miRNP thus formed, under the influence of miRNA and the associated protein(s) achieves its function or follow different fates [37].
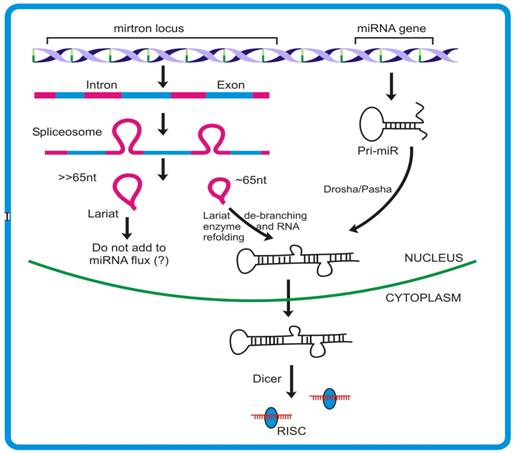
Mirtrons, an emerging concept of microRNA biogenesis: In addition to the canonical miRNA pathway, animals have been shown to follow yet another mode of miRNA biogenesis where intron sequences can produce miRNAs. The hypothesis had emerged through analyses derived from pyro-sequencing of the small RNA pool from Drosophila S2 cell lines where some miR and miR* reads were mapped to the intronic regions. Such miRNAs originating from introns were termed mirtrons [41]. Fourteen mirtrons fromDrosophila and four mirtrons from C. elegans have been identified so far. Given that both protein-coding and non-coding genes possess introns, it is predicted that >80% of miRNA are derived from such sites. Introns denote the region of the transcript, generally flanked by few conserved nucleotide residues, which are removed during RNA processing. The conserved GU-AG along with other sequences brings multiple proteins mediating the removal of the intron from the transcript. The characteristic 2′-5′ phosphodiester bond formed within the intron during splicing result in the formation of lariat like structures. Such structures are acted upon by lariat de-branching enzyme to release the single stranded RNA that is consequently degraded by various nucleases present. However, intron sequences having potential to form hairpin like structures might recruit proteins of miRNA pathway viz.,Dcr1 [40]. Mirtron generation deviates from the canonical miRNA pathway mainly in the non-requirement of DROSHA/DGCR8 proteins that remove the sequences flanking stem region of pri-miR. The unavailability of such sequences leads mirtronic pri-miRs to bypass the DROSHA step. This finding is contrary to the previous observation where Drosha was shown to act on intron prior to its splicing. The knock down experiments with different transcripts involved in canonical miRNA biogenesis viz. Dicer1, Loquacious and Argonaute1, clearly supports that mirtron pathway do not require these proteins for their precursor generation. Moreover, the observation that Dcr-2 and RDR-2 (proteins participating in siRNA generation) knock down do not influence the expression of mirtrons, further ruled out any connection existing between the mirtron and siRNA biogenesis. The expression studies demonstrated that such introns can independently give rise to mature miRNAs, as is the case with miRNA genes. Moreover, like miRNAs, mirtrons also require AGO1 for maturation and follow cell-type specific expression.
The splicing rules guided by the protein complex spliceosome, mediates appropriate release of the structure, termed lariat. These lariats are subsequently effected by another enzyme, called lariat de-branching enzyme which provides an opportunity for this intronic RNA to assume a fold-back structure. The 3′-2 nt overhang of a mirtron forms a substrate for Exportin-5 mediated nuclear exit. The pre-mirtrons thus produced then merge into the mainstream pre-miR pool and follows similar pathway as that of canonical miRNA. In spite of the fact that Drosha is required for mirtron generation, the experiments with Drosha knock down did not show complete inhibition of pre-mirtron biogenesis. Combining the models proposed by Okamura et al., Ruby et al. and Kim et al., it appears that probably both the models are functional and contribute independently to the miR flux. Though these studies have revealed an alternative pathway for miRNA generation, the underlying intrinsic details of the mechanism, e.g., whether large lariat-turned-miR generating RNA can be substrate for DROSHA/PASHA, are yet to be unraveled. Interestingly, though the mirtrons have not yet been identified in plants and other organisms, the presence of introns of pre-miR length directs to the possibility of similar pathway being operative. Since the introns are not subjected to selective pressure it is logical to assume that they are unlikely to preserve their sequences. In light of the above, the mirtron concept might explain the species specificity alongwith the rapid evolution that is observed in miRs. Why an alternate pathway for increasing miR flux has been specifically evolved, poses another interesting puzzle to solve [45]. MicroRNA functionsMicroRNAs show high tissue-specific and temporal expression and are believed to have evolved to take intensive care of developmental pathways that can be achieved by translation suppression (occurring mainly in animals) or target cleavage (occurring mainly in plants). However, there are exceptions to the general functions assigned to animal and plant miRNAs. For example, in animals, miR-196 governs the cleavage of HOXB transcript, and in plants, e.g. Apetala, a transcription factor, is translationally repressed by miR-172. Recently microRNAs have been elucidated to play critical role in conferring immunity to both animals and plants. The target-miRNA recognition, like in case of siRNA, is initiated by seed region sequence. In animals however, the target transcript may possess more than one miRNA recognition site, allowing some miRNAs to bind target at multiple locations in proximity. This probably enhances the silencing effect in a cumulative manner and also confers redundancy to the phenomena, thereby making it more stringent [42]. Plants miRs, on the other hand, have single target binding sites through which they achieve the target fate. However, complex relationship exist between plants miRs and their targets as most of the target transcripts falls under the category of transcription factors and can thus regulate many downstream processes [43]. Interestingly, miRNAs are found to negatively regulate the expression levels of prime RNAi enzymes, viz. dicer and Argonaute. This adds another layer of intricacy to the regulatory network achieved by these molecules [45]. In animals, studies suggest that miRNA binding promotes either deadenylation or decapping of the target which is probably achieved by interaction of RISC associated proteins with cap or poly-A tail associated proteins [43]. However, questions like how miRNA binding employs altogether different mechanisms (translational suppression or activation) is poorly understood. Recent studies have elucidated that miRNAs can bring about translational activation and a probable mechanism has been proposed. Previous studies by Pillai et al. had hinted on the possible cross- talk existing between RNAi and translation activation machinery. These authors showed that AGO2 binding, which is guided by miRNAs (let7 in this case), can relieve the translation arrest of corresponding messages. Lending strong support to these findings, two back-to-back publications from same lab have shed light on our understanding of miRNA and associated proteins in exerting target translation upregulation. The observation that expression of TNF-α, a clinically important protein, vary significantly under starved and nutrient sufficient conditions allowed these authors to hypothesize on the involvement of ARE elements on the mRNA in its stabilization or directly activating translation from the mRNA. Interestingly, the authors found that these AREs bear sequences complementary to miRNAs. Further investigation revealed that two proteins, AGO2 and FXR1 associates, under guidance of miRNAs land onto these sites and exclusively during starved (low glucose) conditions. Importantly, the authors observed similar phenomenon with other miRNAs as well, that led them to hypothesize that miRNAs generally act as repressors during active cellular growth while they tend to behave like translation activator when the conditions are limiting for cell growth. These studies demonstrating the association of miRNAs in translation activation of target mRNAs have provided an altogether new dimension to the functional attributes of miRNAs. Moreover, as against normal localization, microRNAs have been demonstrated to be channeled back to the nucleus in a sequence-dependent manner. This phenomenon allows miRNAs to extend their regulatory roots into other territory, namely the nucleus, thereby providing them an opportunity either to target many under-processed transcripts or bringing about silencing of genomic region(s). Interestingly, this was speculated by Bao et al. where they studied Arabidopsis PHB and PHV mutants that govern the organ polarity and are regulated by miR-165 through degradation pathway. The authors observed that mutants resistant to miR degradation had significantly reduced extent of methylation when compared to wild plants and that there was no impact of other RNAi machinery proteins as a consequence. Studies from Hwang et al. and Bao et al., although with different systems, clearly support the notion that miRNAs play role(s) in RNAi mediated gene silencing. In plants, the miRNA expression pattern is believed to be driven by multiple interacting feedback loops that involve various phytohormones, in particular auxins and gibberellins. Phytohormones regulate transcription of various genes by binding to cis elements and these transcripts possess sites for certain miRNAs. In contrast, transcription of some miRNAs is directly regulated by phytohormones. Such intricate tuning between miR and phytohormones is central to several biological processes [45]. Courtesy International Journal of Biological Sciences [45]
Downloads | Glossary | References | Contact © 2012, Saie Mogre. All Rights Reserved. |