|
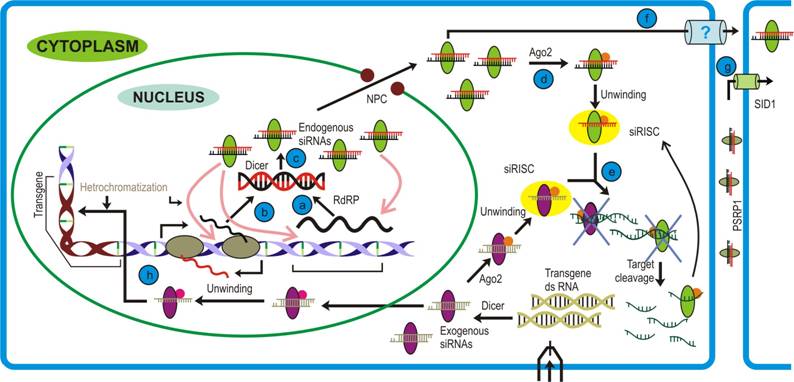
Short-interfering RNAs (siRNAs) are ~20-24 nt long regulatory molecules that besides protecting cell from intrusion of any exogenous nucleic acid (like viruses), are involved in maintaining genome integrity by silencing transcription from undesired loci (retrotransposon, repeat sequences) [23]. siRNA biogenesisThe foremost requirement for siRNA generation is a long double-stranded (ds) RNA molecule. These dsRNAs are formed from any transcription event generating messages with complementary sequences or by some enzymatic activity capable of converting RNA from single strand to double strand (Figure 1, steps a, b). The prime molecule in siRNA generation is Dicer, an RNaseIII type endonuclease. Animals usually encode a single type of Dicer to generate various classes of small RNAs with exceptions of Drosophila and C. elegans each encoding two dicers. Plants on the other hand require multiple dicers. For example, Arabidopsis and Rice possess four and six different Dicer Like proteins (DCLs), respectively. In Arabidopsis, DCL 2, 3, 4 are involved in the generation of different siRNA species while DCL1 is solely responsible for miRNA biogenesis [13]. The activity of each dicer produces siRNAs of characteristic length, e.g. DCL2, DCL3 and DCL4 generate 22, 24 and 21 nt long siRNA species, respectively. Studies carried on Dicer mutants, in both plants and fungi, have allowed us to conclude that these proteins are functionally redundant. The functional redundancy among dicers has been studied in finer details in Arabidopsis, where different combinations of dicer mutants were analyzed on the basis of the size of siRNAs. The siRNA species associated with specific dicer were detected even when the parent dicer was debilitated by mutation. In such a case other dicer(s) may take over the function of the parental dicer, albeit with reduced activity. For instance, in Arabidopsis, DCL1 can produce 21 nt long tasi- RNAs in dcl2dcl3dcl4 mutant and DCL2 can generate tasi-RNAs in dcl4 mutant. Another interesting outcome of these mutant studies was the observation of the existence of functional hierarchy among dicers. This was exemplified by the work on dcl2 and dcl4 mutants, both capable of generating viral siRNAs. However, Arabidopsis dcl4mutant analysis, where viral siRNA accumulation was observed although to a lesser extent, clearly demonstrated that the efficacy of DCL4 to generate viral siRNAs was significantly higher compared to that of DCL2. Analysis of the data available till date strongly suggests that the hierarchy among dicers probably confers stringency to the siRNA mediated pathway [45]. Another important protein involved in siRNA biogenesis in plants, fungi and C. elegans (but not humans) is RNA dependent RNA polymerase (RdRP). The major function of this protein is to generate secondary siRNAs, a step termed signal amplification in siRNA pathway. RdRP can recognize aberrant RNA molecules to produce dsRNAs either in a primer dependent or an independent manner. The dsRNA molecules thus formed are later cleaved by downstream dicer activity. Dicer, in general, possess six domains viz., DExH Helicase, DUF283, PAZ, RNase IIIa, RNase IIIb and RNA Binding Domain. The crystal structures for few individual domains have been solved providing us opportunity to predict model for dicer activity. The role played by each domain is mentioned in the figure along with the expanded form of the abbreviation used. RBD recognizes the duplex RNA structure while the PAZ domain binds to the 3′-2nt overhangs of the cleaved RNA substrate. Dicer has been proposed to act as a single processing centre, where the RBD and the PAZ domains participate in association with RNA molecule and assist both the RNase III domains to come close to form an intra-molecular dimer. These RNase III domains juxtapose in a manner to cleave the ~21 nt duplex RNA molecules from the ds RNA precursors.
The duplex siRNAs are then unwound by helicase activity of Argonaute, a protein recruited by Dicer. This protein has three well characterized domains namely, PAZ, MID and PIWI domains. Both, animals and plants encode multiple Argonautes that participate in different RNAi pathways. Although not all the Argonaute proteins are characterized, the function of few members have been elucidated. For example, AGO2 and AGO1 are prominent members of si- and miRISC, respectively, in the plants and animals. In Arabidopsis, AGO4 plays a crucial role in generating rasi-RNAs that participate in RNA-directed DNA methylation, AGO6 is involved in DNA methylation and transcriptional gene silencing while AGO7 has been shown to participate in both tasi-RNA and long siRNAs biogenesis [42]. Similar to dicer, evidences suggest for the functional redundancy among these proteins. This was observed in Arabidopsis ago1 and ago10 mutant where both were found to share similar phenotypes. Further support came from the studies demonstrating AGO10 performing similar function in ago1 mutant. Here, AGO10 was shown to cleave thePHB transcripts under miR165 guidance and thereby establishing the leaf polarity in the ago1 mutants. Together with the accessory proteins, Argonaute senses the thermal stability of duplex siRNA ends and initiates unwinding from the end with relatively lower thermal energy. Of the two strands, one that is retained with the protein complex (siRISC) is called guide strand while the other (passenger strand) is destined to undergo degradation by exonucleases. In Arabidopsis, the single stranded mature siRNAs are methylated by HEN1 activity thereby rendering stability to these. The single stranded small RNA loaded onto effector complex can be considered analogous to the prokaryotic restriction enzymes that act against any foreign nucleic acid. However, unlike the prokaryotic cellular defense mechanisms which are mediated by restriction enzymes, the small RNAs in eukaryotes can regulate even when the foreign DNA is transcribed to RNA, thus providing a molecular basis to the fact that eukaryotes are evolutionary superior to prokaryotes. On attaining the double-stranded conformation the guide strand of si-RISC activates Argonaute, the RNase activity of which acts specifically on the target sequence at the position complementary to 10 and 11 nt counting from the 5′ end of the siRNA [45]. Classes of siRNAsDepending on the nature of loci and biogenesis of dsRNA precursor different versions of siRNAs have been identified: (i) Trans-acting short interfering RNAs (tasiRNA) are ~21 nt long small RNAs that require endogenous transcript as template that are converted to dsRNA by RNA dependent RNA polymerase (RdRP) activity and subsequently requires the downstream activity of DCL4 and AGO7 to generate functional tasiRNAs. Animals like humans, flies etc., which lack RdRP, are devoid of these small RNA species. Tasi-RNAs resemble miRNAs both in size and function and are involved in targeting non-identical mRNAs. It has been demonstrated that miRNA primed transcripts recruit RdRP that consequently generate tasiRNAs, thereby setting an example of small RNAs mediated regulation of other small RNAs. For instance, miR390 binds to and induces the RdRP activity on primary transcripts and convert them to long dsRNA. In Arabidopsis, the six tasiRNA genes are present that target Auxin Response Factors (ARFs) and MYB transcription factor. One of the recently identified tasiRNA locus, TAS4, has been demonstrated to generate siRNA that targets the transcript at a site which is different from the miR828 cleavage site. This indicates towards the possibility of parallel evolution of tasiRNA, miRNA and their common target in plants[28]. In an alternate pathway, RdRP can also act on aberrant transcripts (usually viral transcripts) converting them to dsRNA and this mechanism is likely to be responsible in preventing cell from any erroneous transcription event that might affect cellular integrity. (ii) Repeat-associated short interfering RNAs (rasi-RNAs) are ~24-26 nt long products of DCL3 activity on dsRNAs formed during unchecked transcription event, usually retro-transposon loci. These loci are generally methylated which prevent transcription through such regions. Like tasiRNA, these also require RdRP for amplifying small RNA pool. Rasi-RNAs play important role(s) during gametogenesis in flies, worms and mammals by modulating the chromatin status, and silencing viral transcripts by recruiting histone modifying proteins [29]. (iii) Scan RNA (scn RNA), another type of relatively long (~29 nts) siRNAs have been reported from protozoan Tetrahymena thermophila. This organism exhibits nuclear dimorphism differing by ~15% at the sequence level. During conjugation, scn RNAs derived from micro-nucleus are generated (reproductive nuclei) and eliminate corresponding loci from its own genome while giving birth to macro-nucleus. This phenomenon requires Argonaute like Twi1 protein, and seems to be an ultimate form of RNA interference wherein organism can efficiently utilize small RNA to produce modified versions of genome from the existing ones [29]. (iv) Long siRNAs (lsiRNAs) constitute the more recently introduced class of siRNAs that are 30-40 nt in length and are induced in response to bacterial infection or growth conditions. Discovered in Arabidopsis, the generation of lsiRNAs require DCL1, DCL4 and AGO7 proteins and depend on other established members of both siRNA and miRNA pathway e.g. RDR6, HYL1, HEN1 etc. One of the lsiRNAs targets a protein that confers resistance against bacterial infection. Interestingly, these lsiRNAs unlike other siRNAs are believed to mediate target degradation by a mechanism previously known in animals but not in plants [29]. Systemic nature of silencingsiRNAs are believed to be a primitive form of immune response evoked against any foreign nucleic acid molecule. Therefore, by corollary, they ought to emanate from the production site to confer rapid cellular defense. This hypothesis is supported from the genetic studies carried out in animals, where import of siRNAs into cells has been demonstrated through a membrane protein called Systemic RNA Interference - Defective (SID-1). Their work on the ectopic expression with cells lacking SID1 shed light on the probable mechanism of the phenomenon. To their interest, they found that SID-1 mediated uptake of dsRNAs was length dependent, with larger molecules (~500 bps) being imported at a faster rate than the smaller (~30 bps) molecules. Moreover, observation that siRNA import is not affected by either cold treatment to cells or ATP depletion suggested towards a passive diffusion mode of this uptake. How the SID-1 protein discriminates between siRNA and miRNA while importing, still remains an open question to the researchers. Phloem, a vascular tissue in land plants, has been implicated in the distribution of sugars, nutrients and other biomolecules across the plant. Recently, Yoo et al. provided evidence for another class of molecules that are mobilized through this route i.e. small RNA. Studies with phloem sap of different plant (rich in sap) species like cucurbits, yucca, and lupin revealed the presence of si- and miRNAs. Further they discovered a novel protein, Phloem Small RNA binding protein-1 (PSRP1), which was functionally similar to SID1 in animals and binds to the small RNA species. This was confirmed by studying viral coat protein silenced and non-silenced lines where authors could find accumulation of Coat Protein (CP) siRNAs in the silenced but not in the non-silenced lines. Nonetheless, the significance of miRNA transport across the phloem still remains to be elucidated. However, the PSRP1 protein is not conserved among the plants. Although the systemic nature of silencing is a well-accepted phenomenon, the underlying mechanism is still ill-defined and demands efforts to resolve the differences between the animal and plant proteins reported till date [45]. Functions of siRNAsThe functionality of siRNA is the consequence following its binding to target sequences and this is governed by a critical region within the siRNA sequence called “seed region”. The ribonucleotides encompassing the 2-7 positions (with reference to 5′ end) of siRNA constitute the “seed region” and are critical to confer siRNAs their target specificity [28]. It is through the “seed region” that RISC lands onto, anneals and consequently brings about target cleavage/repression [29]. Since siRNAs bind to the sequences from which they are derived, they are not under any kind of selection pressure. It may be noted that although the seed region is important in target recognition, the complementarity in other region of siRNA is critical during the cleavage event. siRNAs have been involved in almost all possible nucleic acid regulatory pathways like target cleavage, transcriptional gene silencing and DNA elimination. Moreover, siRNAs from plants have been shown to behave functionally similar to animal miRNAs where the siRNA binding rather than the cleavage leads to the decapping of target transcripts. The exonuclease (XRN4) then acts on decapped mRNAs and bring about target cleavage. siRNAs with lesser complementarity has been demonstrated to suppress the targets at translation level [45]. Courtesy International Journal of Biological Sciences [45] Downloads | Glossary | References | Contact © 2012, Saie Mogre. All Rights Reserved. |