|
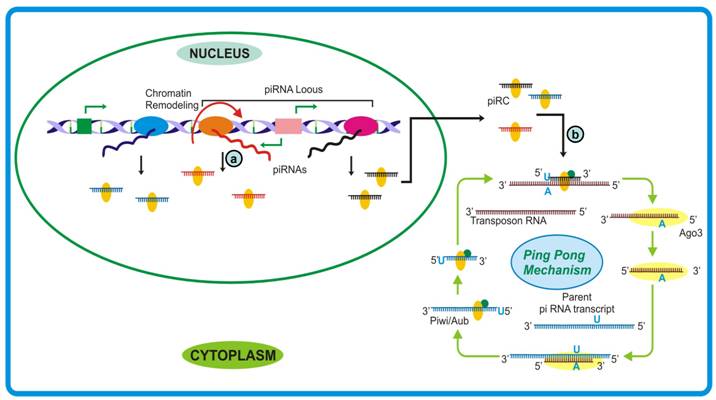
Argonaute family of proteins is a well-established member of executor RNAi complex and is highly conserved amongst various species. Based on amino acid sequence homology, argonaute family has been categorized into two subclades viz., AGO and PIWI (Pelement-induced wimpy testes) [23]. While the AGO members are ubiquitous and associate with both the existing classes of small RNAs, i.e. siRNA and miRNA, the expression of PIWI proteins is restricted to germline cells alone. Genetic studies revealed that mutants for these proteins lead to male sterility suggesting its probable role in spermatogenesis. However, the exact correlation between the over-expression of PIWI and gametogenesis remained unanswered till recently. The independent works of Lau et al., Aravin et al., Grivna et al. and Girard et al. [39] showing that PIWI proteins associate with ~25-31 nt RNA species which are germline-specific have added new dimensions to our knowledge about the varied nature of small RNA world. While Lau et al. and Watanabe et al. focused on purifying the protein complexes followed by cloning of the associated small RNAs, as mentioned above. Cloning and sequencing of the germline small RNA population revealed that majority of the piRNA sequences were mapped to the genomic regions previously thought to be non-transcribed, while others corresponded to intergenic, exonic, intronic and repeat regions. These piRNAs were found to fall in two distinct size categories (24-28 nt and 29-31 nt). These are dispersed throughout the genome and reside in clusters ranging from 1 to 100 kilobases each generating 10 to 4500 piRNAs, eliciting strong bias to either DNA strand. Characterization of the protein complexes revealed the presence of two proteins viz. PIWI and Rec Q1. However, one cannot rule out the possibility of other proteins being loosely bound to the complex which would have subsequently lost during the process. This protein-RNA complex is termed piwi interacting RNA complex (piRC). The presence of RecQ1 as complex constituent was confirmed through its intrinsic ATP-dependent helicase activity [44]. Expression studies have clearly demonstrated that MILI and MIWI (mouse orthologs of PIWI) follow different temporal patterns: while the expression of MILI lasts till pachytene, MIWI expresses till round spermatid stage. Intriguingly, the expression of two different populations of piRNA (24-28 nt and 29-31 nt) follow similar temporal pattern, indicating that each of these proteins might interact with specific piRNA species. piRNA BiogenesisBased on the mapping analysis of piRNA sequences onto genome it is postulated that these piRNA precursors are derived either from the non-overlapping transcripts generated from divergent promoter or from a promoter giving rise to long single stranded RNA. The former postulate was based on the analysis that few piRNA clusters possess gap of few hundred base pairs between transcripts emerging from opposite DNA strands [46]. However, this could not explain the generation of piRNAs from unidirectional promoters [45]. A better understanding of piRNA biogenesis has emerged from the studies conducted by Gunawardane et al. and Brennecke et al. where they proposed a mechanism parallel to secondary siRNA generation, termed as ping-pong model. It was observed that sense piRNAs associate with AGO3 while antisense associate with Piwi/Aub and are complementary till first 10 bases. Also, the 5′ end of antisense piRNA was observed to have strong preference for uridine base and that of sense piRNAs for adenine at position 10. According to the model, the piR-Piwi/Aub complex generated from piRNA cluster binds to target transcript (usually a transposon sequence) and cleaves between 10 and 11 bases. Subsequently, AGO3 binds and guide the cleaved transcript to the piRNA cluster transcript where it follows the hallmark endonucleolytic cleavage after 10th residue (usually adenine). This feed forward loop can rapidly generate sufficient piRNA to take care of any aberrant transcription event especially from retroelements. The cycle can be regulated by sensing reduced production of target transcript and consequently the piRNAs themselves. Whether the piRNA species in an organism are generated following the ping-pong model or by other pathways similar to the biogenesis of si- and miRNAs remains an open question. Moreover, as trigger-target availability is an absolute requirement for the activation of piRNA mediated defense, how these small RNAs take care of recently evolved selfish transposable elements or whether their evolution follows the birth of selfish sequences, still remains obscure [45].
Functional Attributes of piRNAsPrevious studies have shown that PIWI performs multiple functions ranging from epigenetic programming and repression of transposition to post transcriptional regulation. However, in contrast to negative PTGS regulation of si- and miRNAs, piRNAs promote stability of target mRNA and probably enhance the translation as well. Having loci spread throughout the genome, the most important role that could be conferred upon piRNAs would be the patronage of their respective loci. However, in view of the ability of piRC to cleave the cognate transcript, the involvement of piRNA at post-transcriptional level cannot be overruled [45]. Courtesy International Journal of Biological Sciences [45] Downloads | Glossary | References | Contact © 2012, Saie Mogre. All Rights Reserved. |